Capítulo 51.
La continuidad
de la vida I: Desarrollo
Cada vida humana comienza con
la fusión de un espermatozoide y un óvulo . Si el óvulo
recién fecundado de un animal -de cualquier animal- se examina bajo el
microscopio electrónico, no puede encontrarse ningún indicio del
enorme potencial con que está dotada esta célula única.
¿De qué manera las estructuras complejas del embrión ,
y posteriormente del animal adulto, se desarrollan a partir de esta célula
única, aparentemente simple? Esta pregunta es una de las más fundamentales
de la biología y aunque ha concitado la atención de los científicos
durante más de 100 años, aún no se cuenta con una respuesta
completa y definitiva.
En el proceso del desarrollo
, el óvulo fecundado se transforma en un organismo completo, que consiste
en centenares o miles de millones de células y que se asemeja, en gran
medida, a sus progenitores. Este proceso implica crecimiento, diferenciación
y morfogénesis .
El desarrollo en la mayoría
de las especies animales comienza con la fecundación -la fusión
del ovocito y del espermatozoide-. El erizo de mar presenta muchas similitudes
en el proceso de fertilización comparado con los vertebrados y otros
cordados. Los erizos de mar son, desde hace mucho tiempo, los favoritos de los
embriólogos. Una vez formado, el huevo comienza a dividirse por mitosis
. De esta manera, se pone en movimiento la cadena de los fenómenos del
desarrollo.
El desarrollo ocurre en tres
etapas: segmentación , gastrulación y organogénesis . Cuando
la segmentación se completa, el embrión consiste en un cúmulo
de células, la blástula, con una cavidad central, el blastocele.
La segmentación implica una serie de divisiones mitóticas en las
que el volumen citoplasmático del huevo es dividido en numerosas células
nucleadas y más pequeñas, llamadas blastómeras .
La gastrulación da como
resultado el establecimiento de tres capes de tejidos: el endodermo, el mesodermo
y el ectodermo. Cada una de estas capas de tejido primario establecidas origina
tejidos y órganos particulares.
Hacia el final de la gastrulación,
comienzan a aparecer los primeros signos visibles de diferenciación.
En los huevos amniotas , como los de los reptiles, aves y mamíferos,
el embrión, a medida que se desarrolla, forma cuatro membranas extraembrionarias:
el saco vitelino , la alantoides , el corion y el amnios .
Los patrones básicos
de desarrollo son notablemente semejantes en todo el reino animal, incluido
el hombre. Los estudios experimentales sobre invertebrados, anfibios, aves y
mamíferos han provisto de un fundamento sólido a nuestra comprensión
del desarrollo humano.
Fertilización del ovocito
y formación del cigoto
Las primeras etapas del desarrollo
embrionario de un equinodermo son, en varios aspectos importantes, similares
a los que se observan en los vertebrados y otros cordados, lo que indica una
relación evolutiva más íntima que la que se esperaría
sobre la base de la comparación de las formas adultas.
El desarrollo comienza con la
fertilización del óvulo por el espermatozoide. La célula
huevo es de mucho mayor tamaño que el espermatozoide ya que todo el material
necesario para las primeras etapas del desarrollo embrionario está almacenado
en ella. Una vez formado, el huevo comienza a dividirse por mitosis. De esta
manera, se pone en movimiento la cadena de los fenómenos del desarrollo.
En muchas especies, aunque la
activación del óvulo y la mitosis siguen a la fertilización,
también pueden sucederse sin ésta y el embrión puede desarrollarse
por partenogénesis. Los óvulos de muchos invertebrados, y de algunos
vertebrados, son capaces de desarrollarse normalmente en ausencia de espermatozoides
si éstos son activados artificialmente. Los mamíferos, en cambio,
no presentan partenogénesis. Por el contrario, la presencia del pronúcleo
femenino y del masculino son indispensables para el desarrollo normal del embrión.
Segmentación y formación
de la blástula
El embrión comienza a
formar un organismo multicelular mediante un proceso llamado clivaje o segmentación,
proceso que implica una serie de divisiones mitóticas en las que el volumen
citoplasmático del huevo es dividido en numerosas células nucleadas
y más pequeñas. El genoma embrionario transmitido por mitosis
a todas las células no se expresa en los primeros estadios del desarrollo.
Otra característica del clivaje es que, en la mayoría de las especies,
no existe un incremento en el volumen total del embrión durante este
período.
El patrón de clivaje
varía según las especies y está determinado, principalmente,
por la cantidad y la distribución del vitelo en el citoplasma. Cuando
un hemisferio del huevo está relativamente libre de vitelo, las divisiones
celulares ocurren de forma más rápida que en el hemisferio opuesto,
que tiene más vitelo. El hemisferio rico en vitelo se denomina hemisferio
vegetal y el hemisferio opuesto, pobre en vitelo, hemisferio animal. El núcleo
del embrión generalmente se desplaza hacia el hemisferio animal donde
ocurre, en general, el clivaje.
En los embriones con relativamente
poco vitelo -como en el erizo de mar- la segmentación es uniforme, abarca
el embrión entero y forma células de tamaño similar.
El embrión luego pasa
por una etapa de mórula. Luego se crea una cavidad llena de fluido en
el centro del embrión, conocida como blastocele. Cuando
el blastocele está completamente formado, el embrión se llama
blástula y sus células son las blastómeras. La blástula
del erizo de mar tiene, aproximadamente, el mismo tamaño que la célula
huevo a partir de la cual se desarrolló.
En los anfibios, la segmentación
del huevo de los anfibios es diferente de la del erizo de mar principalmente
en el contenido de vitelo. Como los anfibios presentan mayores cantidades de
vitelo, el huevo se divide en forma desigual y se forman células de mayor
tamaño en el hemisferio vegetal. Como ocurre con el huevo del erizo,
la segmentación del huevo de anfibio da como resultado la formación
de una blástula, pero el blastocele del anfibio es pequeño y habitualmente
excéntrico. El blastoporo aparece como una hendidura con forma de media
luna; siempre se forma en el límite entre la media luna gris y el hemisferio
vegetal.
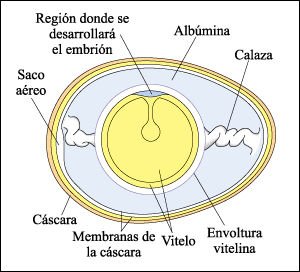
Con respecto a las aves, el huevo de gallina, familiar para todos nosotros, es diferente en muchos aspectos de los huevos mencionados. En primer lugar, está rodeado por una cáscara, que evita la desecación y permite que se desarrolle fuera del agua. En segundo lugar, en relación con su desarrollo terrestre, está rodeado por un sistema de membranas (es un huevo amniota). En tercer lugar, contiene una gran cantidad de vitelo. Esta gran cantidad de alimento almacenado posibilita un período de desarrollo más largo antes de que emerja el animal inmaduro. Como resultado, el pollo, aunque es aún pequeño en el momento del nacimiento, está mucho más adelantado en el desarrollo que la larva plútea de los erizos o el renacuajo.
Anatomía del huevo de pollo
En el momento de su liberación
del ovario en la ovulación, el huevo consiste en la envoltura vitelina
y sus contenidos. La fecundación, si ocurre, tiene lugar en la parte
superior del oviducto. A medida que el huevo se mueve a través del oviducto
se suman las capas de albúmina (la clara de huevo) sintetizadas por las
células del oviducto y liberadas desde ellas. La primera de éstas,
la calaza, es la estructura con forma de cuerda que se observa frecuentemente
cuando se abre un huevo. La calaza mantiene suspendida a la célula huevo
y asegura que el embrión, que se desarrolla a un costado del huevo, esté
siempre hacia arriba, próximo al calor de la madre. Dos membranas de
la cáscara y la propia cáscara se añaden antes de la puesta
del huevo.
El vitelo del huevo de gallina, al igual que el de los huevos de otras aves y también de los reptiles, es tan grande y tan denso que la segmentación no abarca la mayor parte de la masa del huevo. La única parte que se segmenta es una delgada capa de citoplasma, que se sitúa como un casquete en la parte superior del vitelo y contiene al núcleo. La segmentación de esta delgada capa produce una blástula con forma de rombo, conocida como un blastodisco. Las aves, los reptiles y los mamíferos monotremas se desarrollan a partir de un blastodisco.
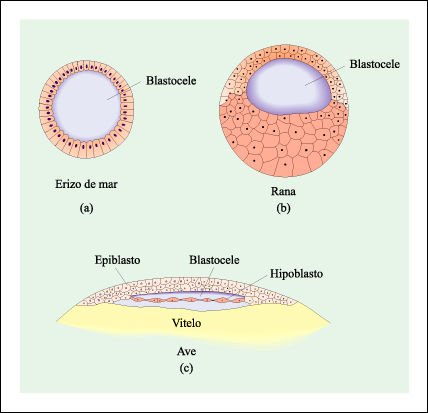
Blástulas de a) un erizo de mar, b) una rana y c) un ave.
En el erizo de mar y en la rana,
la blástula es una esfera de células hueca. En el ave tiene la
forma de un blastodisco -una esfera aplastada de células- que se constituye
en la superficie del vitelo. El blastocele se ubica entre el epiblasto y el
hipoblasto.
Gastrulación y establecimiento
del plan corporal
La formación de la blástula
es seguida por un proceso denominado gastrulación a través del
cual se origina el intestino primitivo y se desarrollan las tres capas de tejido
embrionario: una capa interna, el endodermo, una capa media, el mesodermo y
una capa externa, el ectodermo. Cada una de estas tres capas de tejido primario
origina tejidos y órganos particulares.
En el erizo de mar, la gastrulación
propiamente dicha comienza con la formación del blastoporo, una abertura
en la blástula. Luego, la capa entera de células más próxima
al blastoporo se invagina, moviéndose a través del blastocele
hacia el polo opuesto y formando el arquenterón que finalmente desarrollará
el tubo digestivo. El blastoporo se transformará en el ano. La formación
del ano en el blastoporo -o cerca de él- es la característica
que define a los deuteróstomos, que incluyen a los equinodermos y a los
cordados.
Como resultado de los movimientos que ocurren durante la gastrulación, se forman las tres capas de tejido embrionario mencionadas y se establece el eje anteroposterior del embrión.

Gástrula de erizo de mar.
La gastrulación produce
un embrión de tres capas. El arquenterón se transformará
en el tubo digestivo, y el blastoporo, en el ano. Finalmente, el blastocele
queda casi completamente obliterado. En ésta y en otras ilustraciones
posteriores, el ectodermo es azul, el endodermo es amarillo y el mesodermo es
rojo.
Una vez completada la gastrulación,
se hace evidente que ha ocurrido un proceso de diferenciación celular.
La célula huevo se ha transformado en un número de células
diferenciadas, especializadas, que desempeñan funciones específicas.
La gastrulación en los anfibios sólo difiere de la del erizo de mar en algunos detalles. En Xenopus las células de la blástula tienen diferentes destinos según si estaban en la capa superficial o profunda del embrión. Durante la gastrulación, los tejidos embrionarios primarios -endodermo, mesodermo y ectodermo- se disponen en un patrón de tres capas. Hacia el final de la gastrulación, comienzan a aparecer los primeros signos visibles de diferenciación. El cordamesodermo, una lámina de células mesodérmicas, ha formado la notocorda y el ectodermo neural ha comenzado a engrosarse, formando la placa neural.
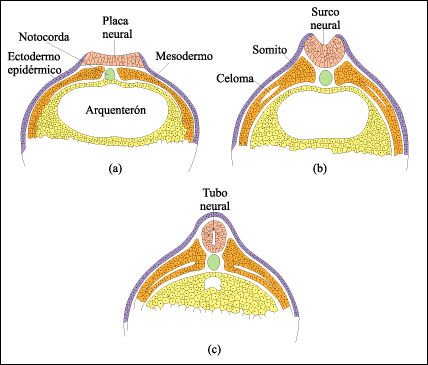
La formación del tubo neural a partir de la placa neural de la rana.
a), b). Las elevaciones engrosadas
del ectodermo neural a derecha e izquierda de la placa neural se curvan formando
el surco neural. Los rebordes del surco neural después se encuentran
y se fusionan. c) Finalmente, el tubo neural resultante se separa del ectodermo
epidérmico.
Simultáneamente, se van
formando los somitos. El celoma se forma entre dos capas de tejido en el mesodermo
de la placa lateral. Quedan así establecidas las principales características
del vertebrado.
En las aves y en los mamíferos, el homólogo del blastoporo es la línea primitiva.

Gastrulación y formación del tubo neural en el pollo.
Uno de los pasos evolutivos
más importantes entre los vertebrados fue el desarrollo del huevo amniota,
que contiene su propia reserva de agua y, por lo tanto, puede ser depositado
en tierra. El agua está contenida dentro de las membranas extraembrionarias.
Estas membranas comienzan como extensiones del blastodisco y cada una está
formada por una combinación de dos de los tipos primarios de tejido.
En los reptiles y en las aves, estas membranas desempeñan papeles esenciales en suministrar al embrión en desarrollo moléculas de alimento y oxígeno, en eliminar productos de desecho nitrogenados y proteger al embrión de la abrasión. En los mamiferos, el saco vitelino es el sitio en el cual las células germinales son retenidas antes de su migración a las gónadas en desarrollo, la alantoides se desarrolla en cordón umbilical, el corion forma estructuras del lado fetal de la placenta, y el amnios, como en aves y reptiles, encierra el embrión en una cavidad llena de fluido.
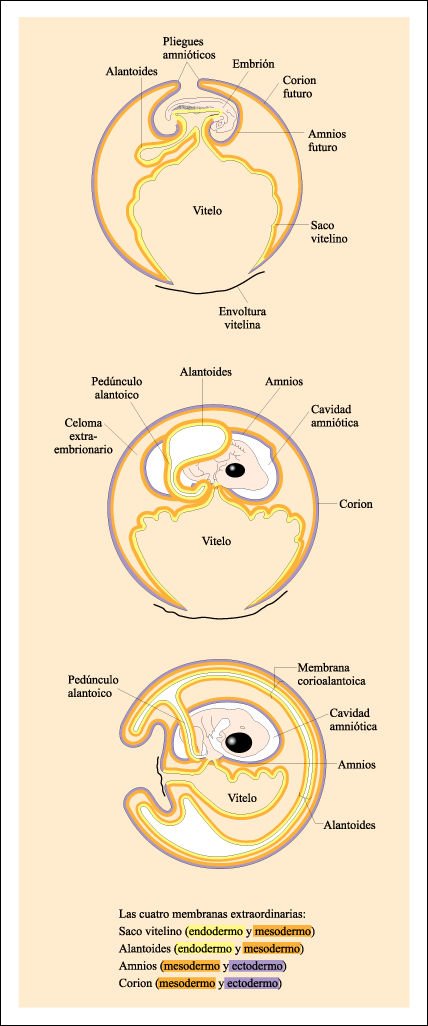
Desarrollo de las membranas extraembrionarias del pollo.
A medida que el embrión
se hace tubular y se separa del vitelo, comienzan a formarse las membranas extraembrionarias.
Una membrana, el saco vitelino, crece alrededor del vitelo y lo rodea casi por
completo. Una segunda membrana, la alantoides, surge como una excrecencia de
la parte posterior del intestino. La tercera y la cuarta se elevan por encima
del embrión, se fusionan y se forman dos membranas separadas. La interna
es el amnios y la externa es el corion. El corion finalmente se fusiona con
la alantoides (membrana corioalantoica), que en etapas posteriores del desarrollo
encierra al embrión, al vitelo y a todas las otras estructuras.
La diferenciación es
el resultado de la expresión diferencial de genes específicos
en el núcleo de una célula. Una variedad de experimentos han demostrado
que, para ciertos tipos celulares, la diferenciación no resulta irreversible
hasta bastante tarde en el proceso del desarrollo. Sin embargo, para muchos
tipos celulares, la potencialidad de desarrollo se ve gradualmente limitada
a medida que la gastrulación avanza y ciertas capas de células
se ubican adecuadamente en el embrión. Este proceso, en el cual queda
fijado el destino de una célula, depende de una serie progresiva de interacciones
entre diferentes tipos de tejidos.
Las últimas etapas del
desarrollo después de la segmentación y de la gastrulación
generalmente se conocen como organogénesis. La organogénesis comienza
con la interacción inductiva entre el ectodermo y el cordamesodermo subyacente.
Cada uno de los tres tejidos primarios formados durante la gastrulación
experimenta luego crecimiento, diferenciación y morfogénesis.
Este proceso es esencialmente el mismo en todos los vertebrados.
Por otra parte, cada sección
del cuerpo tiene una conformación y estructura características.
Sólo unos pocos procesos celulares, repetidos una y otra vez en diversas
permutaciones y combinaciones, parecen ser responsables de la configuración
de las distintas estructuras del cuerpo. Este proceso incluye: 1) incrementos
o decrementos en las tasas de crecimiento y división celular, 2) cambios
en la adhesión de las células a células vecinas; 3) deposición
de materiales extracelulares y 4) cambios en la configuración celular
producidos por extensión o contracción.
Desarrollo del embrión
humano
La fecundación del ovocito
por parte del espermatozoide ocurre habitualmente en el oviducto. Luego, el
huevo o embrión desciende por el oviducto. Simultáneamente experimenta
una serie de divisiones mitóticas que producen un rápido incremento
en el número de células, aunque no en el volumen. Estas células,
las blastómeras, se tornan cada vez más pequeñas con cada
división de segmentación.
En las etapas tempranas, todas
las células son del mismo tamaño, al igual que el erizo de mar,
y son totipotenciales. Sin embargo, las células mantienen su totipotencialidad
durante unas pocas divisiones.
En un principio, el embrión depende exclusivamente del control genético materno y su desarrollo es sostenido por las proteínas, RNA, mitocondrias y otros componentes celulares pertenecientes al ovocito. Esto ocurre hasta que se activa la transcripción en el genoma embrionario. Una vez que el embrión se encuentra en el estadio de mórula, puede ingresar en el útero.
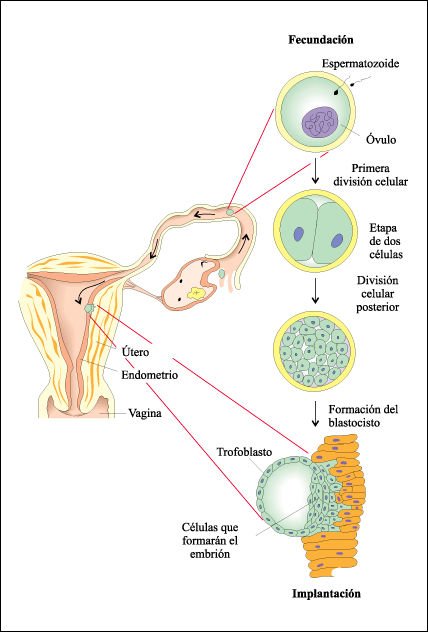
Etapas del desarrollo embrionario humano desde la fecundación hasta la implantación en la pared del endometrio.
El diminuto embrión que
ha alcanzado la etapa de blastocisto invade el endometrio. Una vez realizada
la implantación, comienza a formarse la placenta.
Por un proceso de compactación
se diferencian dos tipos de grupos celulares, uno de los cuales formará
el trofoblasto. Las células trofoblásticas no son capaces de producir
ninguna célula del embrión propiamente dicho, pero son necesarias
para la implantación del embrión en la pared uterina. Las células
descendientes de las células internas de la mórula generarán
la masa celular interna, la cual dará origen al embrión. Así,
la distinción entre blastómeras del trofoblasto y de la masa celular
interna representa la primera diferenciación celular en el desarrollo
de mamíferos.
La mórula adquiere luego
una cavidad interna, el blastocele. La masa celular interna se posiciona sobre
un lado del anillo de células trofoblásticas y esta estructura,
el blastocisto, es bastante diferente de las examinadas hasta ahora. El trofoblasto
es el precursor del corion. Cuando el embrión alcanza el útero,
sale a través de la zona pelúcida y así puede adherirse
a la pared uterina, durante el día 6 del desarrollo. Alrededor de 2 o
3 días después que el embrión llega al útero, el
trofoblasto hace contacto con el epitelio uterino. El embrión humano
es endocrinológicamente activo antes de la implantación; produce
estrógenos -que tienen un efecto local sobre el endometrio- y gonadotrofina
coriónica humana (HCG), la cual estimula al cuerpo lúteo y éste,
así, continúa la producción de estrógenos y progesterona.
Esto impide la menstruación y protege, de esta manera al embarazo.
En la implantación el
embrión penetra en los tejidos del endometrio y es rodeado por vasos
sanguíneos rotos y por la sangre llena de nutrientes que escapa de ellos;
en este momento, la sangre materna entra en contacto directo con el trofoblasto
embrionario.
Al implantarse el embrión,
comienzan a desarrollarse las membranas extraembrionarias que tienen interesantes
similitudes y diferencias con las membranas presentes en el desarrollo de aves
y reptiles. En primer lugar, el saco vitelino no tiene vitelo. Se forma la cavidad
amniótica que forma la segunda membrana extraembrionaria, el amnios.
Como en el pollo, la cavidad amniótica está llena con el líquido
amniótico y así, el embrión se desarrolla en un medio acuoso.
La tercera membrana es el corion, una combinación de células del
trofoblasto y del mesodermo extraembrionario que crece a partir del propio embrión.
El corion representa la porción embrionaria de la placenta y permite
al feto tomar oxígeno y nutrientes de la madre. También es capaz
de secretar hormonas que ayudan al útero materno a retener el embrión
y de producir reguladores de la respuesta inmune que evitan el rechazo materno
del embrión. Alrededor del decimocuarto día, comienzan a formarse
la placenta madura.
En los seres humanos y otros
mamíferos, la alantoides se origina el saco vitelino. En estos organismos,
los desechos metabólicos son transportados en forma de urea y amoníaco
al torrente sanguíneo materno a diferencia de lo que ocurre en el pollo,
en el que son almacenados en forma de ácido úrico.
En los mamíferos, la implantación del embrión y el desarrollo de la placenta son requisitos esenciales para el desarrollo fisiológico normal del feto. Las vellosidades coriónicas otorgan una enorme superficie de intercambio. La placenta se forma como resultado de las interacciones de un tejido materno -el endometrio- con el corion extraembrionario, y está ricamente irrigada por ambos. Sin embargo, los sistemas circulatorios extraembrionarios y materno no están conectados de manera directa, de modo que las células sanguíneas de la madre y del embrión no se mezclan.
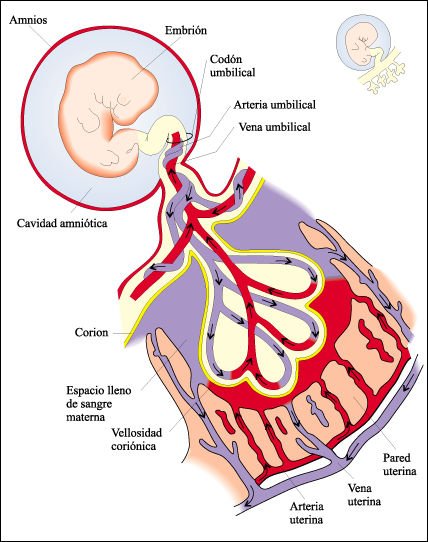
Vellosidades coriónicas.
Desde la placenta, se proyectan
numerosas vellosidades coriónicas digitiformes al espacio de la sangre
materna en la pared del útero. La sangre que llena estos espacios de
la placenta procede de ramificaciones de la arteria uterina.
A través de la delgada
barrera que separa la sangre materna de la fetal, ocurre intercambio de diversas
sustancias: nutrientes solubles, oxígeno, agua y sales pasan a la vena
umbilical desde la sangre de la madre, el dióxido de carbono y los desechos
nitrogenados, llevados a la placenta por las arterias umbilicales, pasan a la
sangre de la madre. Algunas sustancias tóxicas atraviesan fácilmente
la placenta y también lo hacen algunas drogas. La permeabilidad de la
placenta a diferentes sustancias depende del peso molecular de esas sustancias.
Aunque la placenta teóricamente previene el pasaje de microorganismos
desde la madre al feto, algunos patógenos pueden provocar en el feto
enfermedades graves. Los virus atraviesan fácilmente la placenta y también
pueden causar enfermedades severas en el feto o embrión. Así,
la placenta es el órgano excretor del embrión, y es, asimismo,
su superficie respiratoria y su fuente de nutrición.
Cuando el embrión humano
tiene aproximadamente dos semanas, se forma una linea primitiva, seguida por
el desarrollo de una placa neural y un surco neural, que se pliega formando
el tubo neural. Aunque el embrión es aún muy pequeño, la
mayoría de los órganos principales han comenzado a formarse en
estas semanas muy tempranas.
Hacia el final del segundo mes, el embrión, llamado ahora feto, tiene
aspecto casi humano, aunque solamente pesa aproximadamente 1 gramo.
Hacia el
final del tercer mes, todos los sistemas de órganos se han constituido.
Durante el segundo trimestre continúa el desarrollo de los sistemas de órganos, y durante el trimestre final hay un gran incremento en el tamaño y en el peso. El nacimiento ocurre, en promedio, 266 días después de la fecundación.

Un feto humano, poco antes del nacimiento, mostrando las membranas protectoras y los tejidos uterinos que las rodean.
El tapón cervical está
compuesto principalmente de moco. Se desarrolla por influencia de la progesterona
y sirve para mantener a las bacterias y otros agentes infecciosos fuera del
útero. En el 95% de todos los nacimientos, el feto se encuentra con la
cabeza hacia abajo.
El parto se divide en tres etapas:
la dilatación, la expulsión y la etapa placentaria. La dilatación
comienza con el inicio de contracciones del útero y finaliza con la dilatación
completa o apertura del cuello del útero. En esta etapa habitualmente
ocurre la ruptura del saco amniótico (también llamado "bolsa")
con la expulsión de fluidos.
La segunda etapa o etapa de
expulsión comienza con la dilatación completo del cuello y la
aparición de la cabeza del bebé en el cuello del útero.
La tercera etapa, o etapa placentaria, comienza inmediatamente después
del nacimiento del bebé. También implica contracciones del útero
y la expulsión del fluido, de sangre y finalmente de placenta con el
cordón umbilical unido. Esta etapa también es llamada posnacimiento.
El bebé emerge desde
el encierro cálido y protector en el que había estado nutrido
y pudo crecer durante 9 meses. El cordón umbilical -hasta ese momento
su cuerda salvavidas- es cortado inmediatamente después del parto. El
bebé llora con su primer aliento, comienza a respirar regularmente y,
así, se inicia su existencia independiente.
Epílogo
¿Cuándo comenzó
esta vida humana particular? ¿Cuando el espermatozoide se encontró
con el ovocito? ¿Cuando el embrión se transformó en un
feto, visiblemente humano? ¿Cuando el bebé se hizo viable como
una entidad independiente? En el pasado, estos temas se discutían entre
filósofos y teólogos preocupados por la cuestión relativa
al momento en que el alma entraba en el cuerpo. Estos temas han sido revividos
en las controversias éticas y legales relacionadas con el aborto. Sin
embargo, en un sentido evolutivo, ninguno de estos hechos marca el comienzo
de la vida. La vida comenzó hace más de 3.000 millones de años
y fue transmitida desde entonces de organismo a organismo, generación
tras generación, hasta el presente, y se proyecta hacia el futuro, más
allá de lo que la mente humana pueda avisorar. Cada nuevo organismo es,
así, un participante temporal en el hilo continuo de la vida. Del mismo
modo lo es cada espermatozoide, cada ovocito y de hecho -en cierto sentido-
cada célula viva. Sin embargo, cada individuo es una mezcla única
de herencia y experiencia que jamás podrá repetirse y que, por
lo tanto, es irreemplazable. Pero, desde la perspectiva de la continuidad biológica,
una vida humana no dura más que el pestañeo de un ojo.


