Capítulo 38.
Regulación
del crecimiento y desarrollo de las plantas
Las plantas responden a los estímulos de sus ambientes internos y externos.
Estas respuestas les permiten desarrollarse normalmente y mantenerse en contacto
con las condiciones cambiantes que imperan en el medio en que viven.
Las hormonas son factores importantes
en las respuestas de las plantas. Una hormona es un producto químico
producido en tejidos particulares de un organismo y llevado a otros tejidos
de ese organismo, donde ejerce una o más influencias específicas.
Característicamente, una hormona es activa en cantidades extremadamente
pequeñas.
Nuestro conocimiento de las
hormonas vegetales y de sus efectos sobre el crecimiento comenzó con
el estudio del fototropismo. Éste sigue siendo un punto de partida apropiado
en la consideración de las respuestas de las plantas. Los cinco grupos
principales de hormonas que han sido aisladas de las plantas son las auxinas,
citocininas, etileno, ácido abscísico y giberelinas. También
pueden estar presentes otras sustancias reguladoras del crecimiento.
Las plantas responden a diversos
estímulos ambientales. El fototropismo -la curvatura de una planta hacia
la luz- y el geotropismo -la capacidad del vástago para crecer hacia
arriba y de la raíz para crecer hacia abajo- son dos respuestas que otorgan
un alto valor de supervivencia a las plantas jóvenes. En muchas regiones
de la biosfera, los cambios ambientales más importantes que afectan a
las plantas (y, de hecho, a los organismos terrestres en general) son los que
resultan del cambio de las estaciones. La respuesta de los organismos a cambios
en la duración relativa de los períodos de luz y oscuridad en
un ciclo de 24 horas se denomina fotoperiodicidad. Esta respuesta controla la
iniciación de la floración en muchas plantas.
Los ciclos regulares de crecimiento
y actividad que ocurren aproximadamente cada 24 horas se denominan ritmos circadianos.
Estos ritmos son controlados por un oscilador endógeno -el reloj biológico
-. La principal función del reloj biológico, aparentemente, es
suministrar el mecanismo de medición del tiempo necesario para los fenómenos
de fotoperiodicidad.
Algunas especies vegetales muestran
movimientos específicos, rápidos, que se producen como respuesta
al tacto. Además, todas las plantas vasculares parecen responder a otros
estímulos mecánicos con patrones de crecimiento alterados, lo
que da como resultado plantas más bajas y robustas. Muchas angiospermas
también pueden liberar sustancias volátiles, lo que constituye
una comunicación química con otros individuos de la misma especie.
Fototropismo y el descubrimiento
de las hormonas vegetales
Un tropismo es una respuesta
de crecimiento que implica la curvatura de una parte de la planta en el mismo
sentido -o en sentido contrario- en el que actúa un estímulo.
Si la parte de la planta se curva hacia el estímulo, se dice que el tropismo
es positivo, si se curva en sentido contrario, el tropismo es negativo. Una
de las respuestas más evidentes y de utilidad en las plantas es su fototropismo
positivo, o sea, su curvatura hacia la luz.
Charles Darwin y su hijo Francis
realizaron, en 1880, algunos de los primeros experimentos sobre fototropismo.
Trabajando con plántulas de gramíneas, fueron capaces de determinar
que ciertas zonas de las plantas son las que perciben el estímulo necesario
para que ocurra la curvatura hacia la luz. En efecto, ellos observaron que la
curvatura ocurre por debajo del ápice, en una parte inferior del coleóptilo.
La luz que incide en un coleóptilo
en crecimiento hace que se curve hacia ella. La colocación de una cubierta
opaca sobre el extremo de la plántula inhibe su curvatura, pero un collar
opaco ubicado por debajo del ápice no la inhibe. Estos experimentos indican
que "algo" producido en el ápice de la plántula, y transmitido
hacia abajo en el tallo, provoca la curvatura.
En 1926, el fisiólogo
botánico holandés Frits W. Went logró, por medio de un
experimento, separar esa "influencia" de las plantas que la producían.
Went cortó los ápices de los coleóptilos de plántulas
de avena y los colocó sobre una capa de agar, con las superficies de
corte en contacto con el agar, de modo que la hipotética sustancia que
transmitía el estímulo pudiera difundir hacia el agar. Después
de aproximadamente 1 hora, sacó el ápice del copleóptilo
del agar y cortó el agar en pequeños bloques. Luego, colocó
cada bloque sobre el borde de una plántula cortada como se ve en la figura,
que había sido mantenida en la oscuridad. La plántula se curvó
hacia el lado contrario a aquel en el cual había sido colocado el bloque
de agar. La curvatura es el resultado de la influencia de un estímulo,
la auxina, que se desplaza desde el bloque hacia la zona subapical donde promueve
el alargamiento celular. Los bloques de agar que no habían estado previamente
en contacto con el ápice de un coleóptilo iluminado no produjeron
ninguna curvatura o sólo lo hicieron levemente hacia el lado en el que
se había colocado el bloque. Los bloques de agar que se habían
colocado sobre un corte de coleóptilo realizado más abajo no produjeron
efecto fisiológico. Went interpretó los resultados de sus experimentos
como la demostración de que el ápice del coleóptilo ejercía
sus efectos por medio de un estímulo químico y no por un estímulo
físico como, por ejemplo, un impulso eléctrico. Este estímulo
químico fue denominado auxina, vocablo acuñado por Went a partir
de la palabra griega auxein que significa "incrementar". La auxina
fue una de las primeras hormonas vegetales descubiertas.Se sabe ahora que el
fototropismo resulta del hecho de que, bajo la influencia de la luz, una hormona,
la auxina migra del lado iluminado al lado oscuro del ápice caulinar.
Las células del lado oscuro, que contienen más auxina, se alargan
más rápidamente que las del lado iluminado, haciendo que la planta
se curve hacia la luz. Esta respuesta otorga, a las plantas jóvenes,
un alto valor de supervivencia. Sólo la luz azul -o sea la luz de menos
de 500 nanómetros de longitud de onda- resulta efectiva.
Hormonas y la regulación
del crecimiento vegetal
Las hormonas integran el crecimiento,
desarrollo y actividades metabólicas en los distintos tejidos de una
planta y son típicamente activas en cantidades muy pequeñas.
Las hormonas se consideran reguladores
químicos. La respuesta a un "mensaje" regulador determinado
depende no sólo de sus características (o sea de su estructura
química) sino también de la identidad del tejido específico
u órgano blanco sobre el que actúa y de cuándo y cómo
se recibe.Además,
la respuesta a cualquier hormona dada es influida por una variedad de otros
factores del ambiente interno del organismo, entre los cuales cabe destacar
el efecto conjunto de otras hormonas.
Se conocen cinco tipos principales
de hormonas vegetales, o reguladores del crecimiento: las auxinas, las citocininas,
el etileno, el ácido abscísico y las giberelinas. La evidencia
reciente sugiere que otros compuestos también funcionan como hormonas
vegetales.
Las auxinas -ácido indolacético
o AIA- son producidas principalmente en tejidos que se dividen rápidamente,
como los meristemas apicales.
Participan en muchas respuestas
de las plantas, de las cuales la respuesta fototrópica es solo un ejemplo.
Las auxinas provocan el alargamiento
del vástago, promoviendo principalmente el alargamiento celular. En conjunción
con la citocinina y el etileno, las auxinas parecen intervenir en la dominancia
apical, en la cual se inhibe el crecimiento de las yemas axilares, restringiendo
así el crecimiento al ápice de la planta. En concentraciones bajas,
las auxinas promueven el crecimiento de las raíces secundarias y de las
raíces adventicias. En concentraciones más altas, inhiben el crecimiento
del sistema principal de raíces. En los frutos en desarrollo, las auxinas
producidas por las semillas estimulan el crecimiento de la pared del ovario.
La producción disminuida de auxinas se correlaciona con la abscisión
de frutos y hojas. La capacidad de las auxinas para producir estos variados
efectos parece resultar de las diferentes respuestas de los distintos tejidos
"blanco" y de la presencia de otros factores, incluyendo otras hormonas.
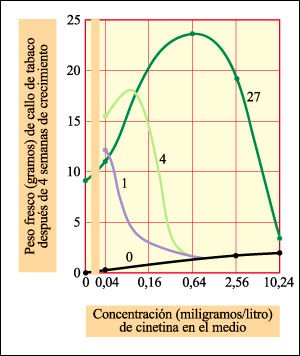
La citocininas promueven la división celular. Alterando las concentraciones relativas de auxinas y citocininas, es posible cambiar los patrones de crecimiento de un tejido vegetal indiferenciado.
La respuesta de las células de tabaco en cultivo de tejido a combinaciones de auxina (AIA) y citocinina (cinetina). Las concentraciones de AIA (en miligramos por litro) se indican en las curvas.
La cinetina sola tiene poco efecto en el crecimiento del tejido indiferenciado
de tabaco (un callo). El AIA sólo hace que el cultivo crezca hasta un
peso de aproximadamente 10 gramos, independientemente de la concentración
usada. Cuando ambas hormonas están presentes, el crecimiento se incrementa
en gran medida. Nótese, sin embargo, que cuando se exceden las concentraciones
óptimas, la tasa de crecimiento declina.
El etileno es un gas producido
por los frutos durante el proceso de maduración, proceso que ese mismo
gas promueve. Desempeña un papel central en la abscisión de las
hojas y se piensa que es un efector de la dominancia apical. El ácido
abscísico, una hormona inhibidora del crecimiento, puede estar involucrado
en la inducción de la dormición en las yemas vegetativas y en
el mantenimiento de la dormición de las semillas.
Las giberelinas estimulan el alargamiento del vástago, inducen el repentino crecimiento y floración de muchas plantas y también están implicadas en el crecimiento del embrión y de la plántula. En las gramíneas estimulan la producción de enzimas hidrolíticas que actúan sobre el almidón almacenado, los lípidos y las proteínas del endosperma, convirtiéndolos en azúcares, ácidos grasos y aminoácidos que nutren a la plántula.
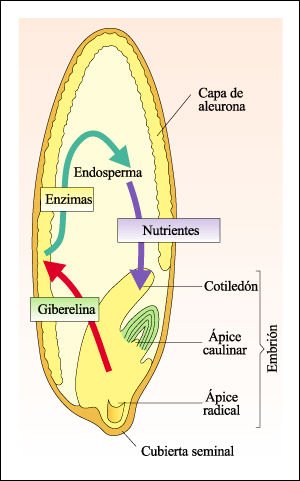
La acción de la giberelina en una semilla de cebada.
El embrión libera giberelina que difunde hacia la capa aleurónica.
La giberelina hace que las células de aleurona sinteticen enzimas que
digieren las reservas alimenticias del endosperma convirtiéndolas en
moléculas más pequeñas. Estas moléculas son transportadas
a las regiones del embrión en crecimiento.

Geotropismo
El fototropismo otorga un alto
valor de supervivencia a una planta joven. Otra respuesta con un alto valor
de supervivencia es la capacidad de responder a la gravedad, enderezándose,
de modo que el vástago crece hacia arriba y las raíces no estáhacia
abajo. Esta respuesta es conocida como geotropismo; al igual que el fototropismo,
se cree que el geotropismo involucra a las auxinas.
Este proceso, el geotropismo,
implica cuatro pasos secuenciales: la percepción de la gravedad, la producción
de señales en células sensoras de la gravedad, la transducción
de señales tanto dentro de células sensoras como entre células
y, finalmente, la respuesta. Una cuestión central se vincula con el modo
en que se detecta la influencia de la gravedad: ¿cómo "sabe"
una plántula si está en posición horizontal?
Las hormonas y los iones son
solubles por lo que la gravedad misma no debería tener efecto sobre su
distribución. De acuerdo con una hipótesis vigente, la gravedad
es percibida por organelas especializadas conocidas como estatolitos. Se demostró
que los estatolitos se corresponden con los amiloplastos. La acumulación
de almidón en tales estructuras es necesaria en este proceso. Cuando
una raíz crece en forma vertical, los amiloplastos se reúnen cerca
de las paredes inferiores de las células centrales.
Sin embargo, si la raíz
se coloca en posición horizontal, los amiloplastos se deslizan hacia
abajo y se disponen cerca de las que previamente eran paredes orientadas en
forma vertical. A los pocos minutos, la raíz comienza a curvarse hacia
abajo y los amiloplastos retornan gradualmente a su posición original.
Fotoperiodicidad
Las plantas son capaces de acomodarse
a los cambios estacionales en virtud de su capacidad de anticipar el calendario
anual de los acontecimientos: la primera helada, las lluvias de primavera, los
largos períodos de sequía, los largos períodos de crecimiento,
y hasta la época en que florecen las plantas vecinas de la misma especie.
Para muchas plantas, todas estas determinaciones se hacen de la misma manera:
"midiendo" los períodos relativos de luz y oscuridad. Este
fenómeno se conoce como fotoperiodicidad.
Algunas plantas, conocidas como plantas de días largos, florecerán sólo cuando los períodos de luz excedan una longitud crítica. Otras plantas, las de días cortos, florecerán sólo cuando los períodos de oscuridad sean mayores que cierto tiempo crítico. Las plantas neutras dan flores independientemente del fotoperíodo. Los experimentos han mostrado que el período de oscuridad, y no el período de luz, es el factor crítico.
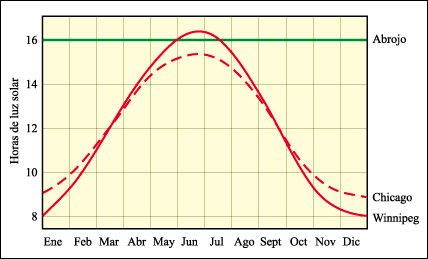
La longitud relativa del día y de la noche determinan cuándo florecerán muchas plantas.
Las dos curvas de la figura anterior muestran el cambio anual de la longitud
del día en dos ciudades de América del Norte, ubicadas a diferentes
latitudes (recuérdese que, en el hemisferio norte, los meses de junio,
julio y agosto corresponden al verano). La línea verde indica el fotoperíodo
que resulta eficaz para el abrojo, que requiere menos de 16 horas de luz para
florecer. En Chicago, el abrojo puede florecer al finalizar el mes de julio,
cuando las plantas ya son adultas pero, en Winnipeg, las yemas florales no aparecen
hasta los primeros días de agosto de modo que, habitualmente, la helada
mata a las plantas antes de que maduren las semillas

Una planta de abrojo.
El abrojo, una planta de día corto que puede soportar toda clase de inclemencias
ambientales, ha sido importante para estudios experimentales de fotoperiodicidad.
Cada receptáculo espinoso es una inflorescencia con dos flores.
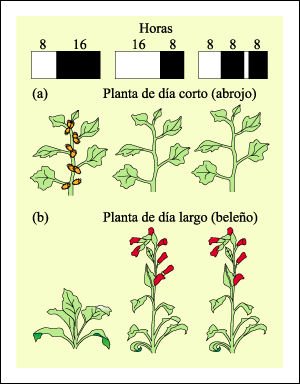
Experimentos de fotoperiodicidad.
Los experimentos de fotoperiodicidad mostraron que las plantas miden la longitud
del período de oscuridad y no del de luz. a) Las plantas de día
corto florecen sólo cuando el período de oscuridad excede cierto
valor crítico. Así, el abrojo, por ejemplo, florece con 8 horas
de luz y 16 horas de oscuridad. Si el período de 16 horas de oscuridad
es interrumpido, aunque sea brevemente, como se muestra a la derecha, la planta
no florece. b) La planta de día largo, por otra parte, que no florece
con 16 horas de oscuridad, lo hace si el período de oscuridad se interrumpe.
Las plantas de día largo florecen sólo cuando el período
de oscuridad es menor que cierto valor crítico.
El fitocromo, un pigmento presente comúnmente en pequeñas cantidades en los tejidos de las plantas, es uno de los sistemas receptores que detectan las transiciones entre la luz y la oscuridad. El pigmento existe en dos formas, Pr y Pfr. Pr absorbe luz roja, mientras que Pfr absorbe luz roja-lejana. Pfr es la forma biológicamente activa del pigmento; entre sus muchos efectos conocidos, promueve la floración en las plantas de días largos, inhibe la floración en las plantas de días cortos, promueve la germinación en las semillas pequeñas de plantas como la lechuga, e interviene en numerosos procesos de los vegetales.
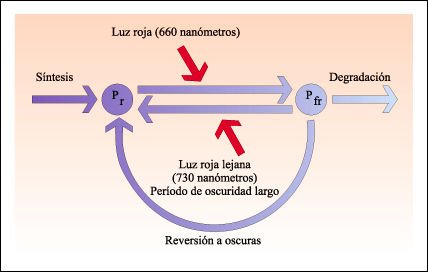
Interconversión de las dos formas del fitocromo.
Los fitocromos son sintetizados en la forma Pr. Pr cambia a Pfr cuando se lo
expone a luz roja. Pfr es la forma activa que induce la respuesta biológica.
Pfr revierte a Pr cuando se lo expone a la luz roja lejana (730nm). En la oscuridad,
Pfr lentamente revierte a Pr o es degradado.
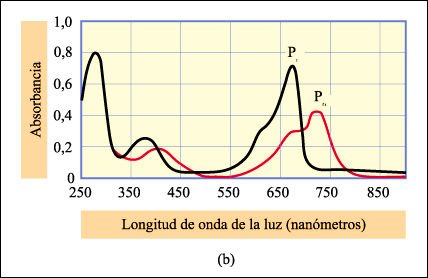
Espectros de absorción de las dos formas del fitocromo.
Las similitudes entre los espectros
de acción de las respuestas biológicas y los espectros de absorción
de los pigmentos suministraron importante evidencia de que el fitocromo es el
pigmento responsable de las respuestas.
Ritmos circadianos y relojes
biológicos
¿Cómo puede una
planta de espinaca distinguir un día de 14 horas de un día de
13,5 horas? La medición del fotoperíodo requiere, por un lado,
de fotorreceptores que permitan distinguir el día de la noche y, por
otro lado, de un mecanismo que mida las horas de oscuridad. En cuanto a los
fotorreceptores, se sabe que tanto los fitocromos como los criptocromos están
involucrados. El tema pendiente es, ¿cómo miden las plantas la
duración del período de oscuridad?
Esta pregunta nos lleva a otro grupo de fenómenos fácilmente observables. Algunas especies de plantas tienen flores que se abren por la mañana y se cierran al atardecer. Otras extienden sus hojas a la luz del Sol y las pliegan hacia el tallo durante la noche.

Hojas de una planta de vinagrillo (oxalis), durante el día a) y la noche b).
Una hipótesis reciente, pero aún sin pruebas, es que los "movimientos
de sueño" evitan que las hojas absorban la luz que refleja la luna
en las noches muy claras, protegiendo los fenómenos fotoperiódicos.
Estos movimientos diurnos continúan
aunque las plantas se mantengan en condiciones lumínicas constantes.
Actividades menos evidentes, tales como la fotosíntesis, la producción
de auxinas y la tasa de inhibición celular, también tienen ritmos
diarios. Los ritmos que continúan con un período cercano a 24
horas aun cuando todas las condiciones del ambiente se mantengan constantes
se llaman ritmos circadianos y se han encontrado en todos los organismos eucarióticos
y en algunos procarióticos.
Si bien la persistencia de los
ritmos circadianos bajo condiciones lumínicas constantes sugiere que
son generados por un oscilador endógeno, durante varios años,
los biólogos debatieron acerca de si estos ritmos podrían estar
determinados por alguna fuerza ambiental poco perceptible, como los rayos cósmicos,
el campo magnético o la rotación de la Tierra.
Virtualmente, todos los biólogos
están de acuerdo ahora en que los ritmos circadianos son endógenos,
o sea, se originan dentro del propio organismo y son controlados por lo que
se conoce como reloj biológico. La evidencia que apoya la idea del reloj
biológico interno es que, en condiciones ambientales constantes, los
ritmos circadianos tienen períodos que no son de 24 horas exactas. Diferentes
especies y diferentes individuos de la misma especie suelen tener ritmos ligeramente
diferentes, pero constantes, a menudo de hasta una o dos horas más largos
o más cortos que 24 horas.
Dado que el período endógeno
de los ritmos no es exactamente igual a 24 horas, en condiciones naturales los
relojes deben ser diariamente ajustados o sincronizados por el ambiente. Los
factores más importantes en la sincronización de los relojes son
las transiciones luz/oscuridad que ocurren al atardecer y al amanecer. Actualmente
se sabe que tanto los fitocromos como los criptocromos son responsables de la
sincronización por luz de los relojes de las plantas.
En los últimos años
se han acumulado evidencias de que el reloj está constituido por proteínas
que regulan negativamente la expresión de sus propios genes, constituyendo
una retroalimentación negativa de 24 horas de duración.
Los relojes biológicos desempeñan un papel importante en muchos
aspectos de la fisiología vegetal y animal, sincronizando acontecimientos
internos y externos.
Respuestas táctiles
El enrollamiento es causado
por las velocidades de crecimiento variables en los diferentes lados del zarcillo.
Este proceso, llamado circumnutación, incrementa las posibilidades de
que los zarcillos encuentren un soporte. Cuando el ápice de un zarcillo
toca cualquier objeto, responde al tacto formando una espiral apretada. Las
células que tocan el soporte se contraen ligeramente y las del lado externo
se alargan. Las células epidérmicas especializadas de los zarcillos
son los sensores al tacto, pero el mecanismo por el cual estas células
inducen el enrollamiento se desconoce.
La evidencia actual indica que
probablemente intervengan las auxinas y el etileno; estas hormonas hacen que
los zarcillos cortados se tuerzan aun en ausencia de tacto.
Muchas plantas responden al tacto, entre ellas, los zarcillos.

Zarcillos de una planta cucurbitácea (Cucumis anguria).
Ciertos estudios han mostrado que los zarcillos pueden almacenar la "memoria"
de la estimulación táctil. Por ejemplo, si los zarcillos del guisante
se mantienen en la oscuridad durante tres días y luego se golpean, no
se enroscan, tal vez porque requieren ATP. Sin embargo, si se los ilumina a
las dos horas después de tocarlos, muestran la respuesta de enrollamiento.
Una respuesta más rápida al tacto ocurre en la planta sensible Mimosa pudica. Unos pocos segundos después de que se toca una hoja, el pecíolo se dobla y los folíolos se pliegan. Esta respuesta es el resultado de un cambio súbito en la turgencia de las "células motoras" especializadas presentes en los engrosamientos articulados llamados pulvinos, los que -a su vez- se encuentran en la base de los folíolos y de las hojas. Estas mismas células intervienen en los "movimientos de sueño", que también ocurren en Mimosa. Dependiendo de qué planta se trate y de la intensidad del estímulo, pueden ser afectadas una sola o todas las hojas de la planta.

Planta de sensitiva (Mimosa pudica).
a. Posición normal de las hojas y los folíolos.
b. Respuesta al tacto. Se ha supuesto que estas reacciones pueden evitar el
marchitamiento (cuando ocurren en respuesta a vientos fuertes), espantar insectos
o desalentar a herbívoros más grandes.
El colapso de los folíolos
es causado por cambios rápidos de turgencia en las células de
los pecíolos. Estos cambios están acompañados por la secreción
de taninos. Los taninos tienen un sabor astringente y repelen a los herbívoros.
Una serie de reacciones parecen
estar involucradas en la transmisión del estímulo. El estímulo
sensorial aparentemente se traduce rápidamente a una señal eléctrica,
semejante a un impulso nervioso en los animales, que pasa a lo largo del pecíolo
y que puede ser detectado colocando microelectrodos en la planta. La señal
eléctrica, a su vez, desencadena una señal química que
hace que la membrana celular de las células motoras se vuelva más
permeable a los iones potasio (K+) y cloruro (Cl-). El movimiento de estos iones
hacia afuera de las células motoras hace que el agua deje las células
por diferencia de potencial hídrico. Como resultado de esta pérdida
de agua, las células motoras se colapsan, causando, por lo tanto, un
movimiento de la hoja o del folíolo.
Las respuestas rápidas
al tacto ocurren también en la captura de presas por parte de las plantas
carnívoras: las atrapamoscas dionea y drosera. Las hojas de la dionea
tienen dos lóbulos articulados, cada uno de los cuales está equipado
con tres pelos sensoriales. Cuando un insecto se posa sobre una hoja, se frota
contra los pelos, generando un impulso eléctrico que provoca el cierre
de la hoja. Los bordes dentados se cierran como una trampa para osos, haciéndolo
en menos de medio segundo. Una vez que el insecto es atrapado, las mitades de
la hoja gradualmente se acercan una a otra y el animal capturado es presionado
contra las glándulas que secretan enzimas digestivas, situadas en la
superficie interna de la trampa.
El impulso eléctrico
generado cuando el insecto se fricciona contra los pelos gatillo activa una
enzima que, usando el ATP para obtener energía, bombea iones H+ hacia
las paredes de las células epidérmicas que se encuentran en la
superficie externa de la bisagra de la trampa.
La expansión celular
rápida e irreversible que resulta de la acidificación de las paredes
celulares cierra la trampa. La magnitud en que la salida de iones H+ afecta
los flujos de entrada y salida de otros iones podría ser -sin embargo-
la que determine tal expansión. Durante este proceso, las células
de la superficie interna de la "bisagra" siguen creciendo a su velocidad
lenta habitual; aproximadamente 10 horas después, su crecimiento supera
el crecimiento de las células de la superficie externa, y la trampa se
abre. Como resultado de ambos procesos de crecimiento, la hoja es ligeramente
más grande que antes de cerrarse.
Ciertos estudios han mostrado que un impulso eléctrico también interviene en el mecanismo por el cual la drosera atrapa insectos.

La atrapamoscas Drosera.
La atrapamoscas Drosera es una planta diminuta que mide sólo entre 2 y 5 centímetros de ancho y tiene tentáculos en forma de garrote en la superficie superior de las hojas. Estos tentáculos secretan un líquido pegajoso que atrae a los insectos. Cuando un tentáculo captura a un insecto, los otros tentáculos se curvan hacia él hasta que el insecto se ahoga. Los tentáculos también secretan enzimas digestivas.

Los nepentes.
Los nepentes son plantas que atraen a los insectos hacia sus hojas tubulares por medio del néctar. Siguiendo al néctar desde el borde de la hoja, el insecto se encuentra con una alfombra de pelos transparentes y finos. Cuando el sendero de néctar finaliza y el insecto quiere rehacer el camino, se topa con las puntas de estos delgados pelos que apuntan hacia abajo y bloquean el camino. Si se mueve hacia el interior, se encuentra en una superficie cérea y resbalosa en la que patina terminando por caer en un caldo maloliente, mezcla de agua de lluvia, enzimas digestivas y bacterias en descomposición, situado en la base.

Otra atrapamoscas, la dionea.
Además de las respuestas especializadas de algunas plantas, el tacto
y otros estímulos mecánicos también pueden tener efectos
amplios en los patrones de crecimiento. Las plantas, en su ambiente natural
están, por supuesto, sujetas a estímulos de este tipo en forma
de viento, gotas de lluvia y movimiento de los animales que pasan.
La dionea tiene hojas que se
cierran alrededor de cualquier insecto que se mueva sobre su superficie. El
cierre de las hojas ocurre un insecto toca uno o dos de los tres "pelos
gatillo" que se encuentran en la parte media de cada lóbulo foliar.
La dieta habitual de la dionea en estado natural consiste usualmente en invertebrados
no voladores
Comunicación química
entre las plantas
Las plantas pueden liberar sustancias
volátiles con las que "se comunican" con otros individuos de
la misma especie, generando en ellos la síntesis de los productos químicos
defensivos antes de que ocurra algún daño real.
En algunas especies, la producción
de compuestos tóxicos se inicia o se incrementa en respuesta al daño
infligido a la planta por herbívoros tales como insectos masticadores
o grandes animales. Las concentraciones más elevadas de estas sustancias
detienen la depredación ulterior y, por lo tanto, protegen a la planta
de un daño aun mas grave.
Estudios recientes han mostrado
que las plantas hacen algo más que protegerse a sí mismas: aparentemente
también "previenen" a las plantas vecinas de la misma especie
para que movilicen sus defensas antes del ataque.


