Capítulo 47.
Integración
y control II: el sistema nervioso
El sistema nervioso, junto con
el sistema endocrino, integra y controla las numerosas funciones que capacitan
a un animal para regular su ambiente interno y reaccionar y enfrentar al ambiente
externo.
Dentro del reino animal se puede
constatar una complejidad sensorial creciente, una mayor capacidad de procesamiento
de la información y una tendencia a la centralización de grupos
neuronales en ganglios. La evolución ha favorecido una especialización
en los sistemas nerviosos en recibir información, codificarla y transmitirla
de neurona en neurona.
El sistema nervioso central
consta del cerebro y la médula espinal, que en los vertebrados, están
contenidos en el cráneo y la columna vertebral. La porción del
sistema nervioso que se encuentra fuera del sistema nervioso central constituye
el llamado sistema nervioso periférico.
En los vertebrados, las neuronas
de salida del sistema nervioso periférico están organizadas en
dos divisiones principales: el sistema nervioso somático y el sistema
nervioso autónomo. El sistema autónomo tienen a su vez dos ramas-
el sistema simpático y el parasimpático- que son anatómica,
fisiológica y funcionalmente distintas. La unidad funcional del sistema
nervioso es la neurona o célula nerviosa. Una neurona está formada
por dendritas que reciben estímulos; un cuerpo celular que contiene el
núcleo y la maquinaria metabólica que también recibe estímulos
y un axón o fibra nerviosa, que envía estímulos a otras
células.
La información recibida
de los ambientes interno y externo, y las instrucciones llevadas hacia los efectores
son transmitidas en el sistema nervioso en forma de señales electroquímicas.
En el estado de reposo, hay una diferencia en carga eléctrica entre el
interior y el exterior de la membrana celular del axón -el potencial
de reposo-. Luego de la estimulación apropiada ocurre un potencial de
acción, que es una inversión transitoria en la polaridad de la
membrana. El potencial de acción que se transmite a lo largo de la membrana
axónica es el impulso nervioso. Como todos los potenciales de acción
tienen la misma amplitud, el mensaje llevado por un cierto axón puede
variar sólo con un cambio en la frecuencia o en el patrón de los
potenciales de acción. En las fibras mielínicas, el impulso nervioso
salta de un nodo a otro de la vaina de mielina, acelerándose así
la conducción.
Las neuronas transmiten señales
a otras neuronas a través de uniones llamadas sinapsis. En la mayoría
de las sinapsis, la señal cruza la hendidura sináptica en forma
de una sustancia química, un neurotransmisor, que se une a un receptor
específico en la membrana de la célula postsináptica.También
intervienen neuromoduladores. La unión de un neurotransmisor o de un
neuromodulador a su receptor puede abrir o cerrar un canal iónico de
membrana o poner en movimiento un segundo mensajero. El efecto final es un cambio
en el voltaje de la membrana de la célula postsináptica.
Una sola neurona puede recibir
señales de muchas sinapsis y, según la suma de las señales
excitadoras e inhibidoras, se iniciará o no un potencial de acción
en su axón. Así, las neuronas individuales funcionan como importantes
centros de transmisión y control en la integración de la información
por el sistema nervioso.
Evolución de los sistemas
nerviosos
Al comparar los sistemas nerviosos
de los invertebrados, desde los más simples hasta los complejos, se evidencia
una tendencia a la concentración de tejido nervioso en zonas especializadas
y protegidas.
En los vertebrados, el sistema nervioso es dorsal, y se encuentra notablemente desarrollado. Sus centros principales de procesamiento -la médula espinal y el cerebro- están encerrados y protegidos por los huesos de la columna vertebral y del cráneo. En la evolución de los vertebrados se observa una tendencia hacia la cefalización. La integración precisa que acompaña a esta centralización posibilita comportamientos complejos.
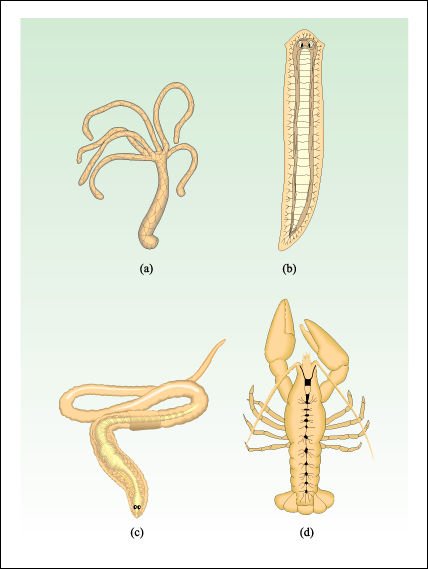
a) Hydra, un cnidario; b) una planaria; c) una la lombriz de tierra. d) un cangrejo de río.
En Hydra a), un cnidario, el impulso nervioso se propaga de modo difuso a lo
largo de la red nerviosa desde el área de estimulación. En la
planaria, b), hay dos cordones nerviosos longitudinales y cierta agregación
de ganglios y órganos sensoriales en el extremo anterior. En los anélidos,
como la lombriz de tierra c), los cordones nerviosos longitudinales están
fusionados en un doble cordón nervioso ventral. En el cangrejo de río
d), un artrópodo, el cordón nervioso también es doble y
ventral, con una serie de ganglios, casi tan grandes como el cerebro, que controlan
segmentos particulares del cuerpo.
Organización del sistema
nervioso de los vertebrados
El sistema nervioso de los vertebrados
tiene subdivisiones que pueden ser distinguidas por criterios anatómicos,
fisiológicos y funcionales. La principal es la subdivisión en
sistema nervioso central -el cerebro y la médula espinal- y sistema nervioso
periférico -las vías sensoriales y motoras que llevan información
hacia y desde el sistema nervioso central-. Las vías motoras se dividen
a su vez en el sistema nervioso somático, con control voluntario sobre
el músculo esquelético y en el sistema nervioso autónomo,
que controla en forma inconsciente al músculo liso, al cardíaco
y a las glándulas. El sistema autónomo, a su vez, se subdivide
en sistema simpático y sistema parasimpático.
La unidad funcional del sistema
nervioso es la neurona, que tiene un cuerpo celular, un axón y frecuentemente
muchas dendritas. Hay cuatro clases de neuronas: neuronas sensoriales; interneuronas;
neuronas de proyección y neuronas motoras. Muchas están rodeadas
y aisladas por células de la glia, llamadas neuroglia en el sistema nervioso
central y células de Schwann en el sistema nervioso periférico.
Las células de la glia,
si bien no participan directamente en la producción del impulso nervioso,
proveen la vaina de mielina que acelera la transmisión de las señales
a través de las neuronas, actúan como tejido de sostén,
facilitan la nutrición de las neuronas y la remoción de sus desechos
metabólicos y sirven como guías para el desarrollo neuronal.
En vertebrados e invertebrados, los cuerpos de las células nerviosas frecuentemente se encuentran agrupados en ganglios si se encuentran a nivel del sistema nervioso periférico y núcleos si están en el sistema nervioso central. Los axones, que constituyen las fibras nerviosas, también se agrupan formando haces: se llaman tractos cuando están en el sistema nervioso central y nervios cuando están en el sistema nervioso periférico.
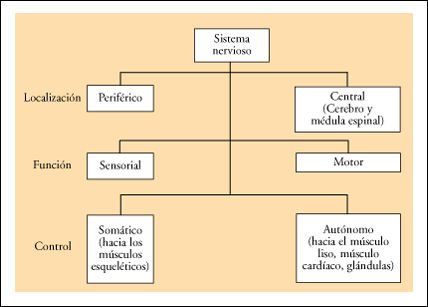
Subdivisiones del sistema nervioso de un vertebrado como el Homo sapiens.
El sistema nervioso de los vertebrados
consiste en un sistema nervioso central -el cerebro y la médula espinal-
y un sistema nervioso periférico -una vasta red de nervios que conectan
el sistema nervioso central con todas las otras partes del cuerpo-. Las neuronas
sensoriales llevan información al sistema nervioso central y las neuronas
motoras la llevan desde ese sistema. Las neuronas motoras están organizadas
en los sistemas somático y autónomo, y el sistema autónomo
contiene dos divisiones: la simpática y la parasimpática.Dentro
del sistema nervioso central, la médula espinal constituye el enlace
entre el cerebro y el resto del cuerpo. Es un cilindro delgado que en un corte
transversal se ve dividido en un área central de materia gris y un área
externa de materia blanca. La materia gris de la médula consiste fundamentalmente
en interneuronas, cuerpos celulares de neuronas motoras y neuroglia. La materia
blanca consiste en tractos de fibras que corren a lo largo de la médula
espinal, formados principalmente por axones.
La médula se continúa
con el tallo cerebral, en la base del cerebro; éste contiene tractos
de fibras que conducen señales hacia y desde la médula espinal
y también los cuerpos celulares de las neuronas cuyos axones inervan
los músculos y las glándulas de la cabeza. Además, dentro
del tallo cerebral hay núcleos que controlan algunas de las funciones
reguladoras automáticas importantes, como el control de la respiración
y de la presión sanguínea.
El sistema nervioso central
se encuentra protegido además por capas de membranas -las meninges- que
regulan el pasaje de sustancias desde la circulación general hacia el
tejido nervioso -la barrera hematoencefálica- y hacia el líquido
cefalorraquídeo -la barrera hematocefalorraquídea-. Las células
gliales que rodean a esos capilares también contribuyen a establecer
una barrera.
Sólo atraviesan las barreras
las sustancias liposolubles y de bajo peso molecular. Existen zonas del sistema
nervioso central que se encuentran por fuera de estas barreras, y que funcionan
como sensores del estado del organismo.
El sistema nervioso periférico
está constituida por neuronas cuyos axones se extienden desde el sistema
nervioso central a los tejidos y órganos del cuerpo. Incluyen tanto a
neuronas motoras eferentes como a neuronas sensoriales, aferentes. Las fibras
de las neuronas motoras y de las neuronas sensoriales están unidas formando
nervios: los nervios craneales y los nervios espinales. Pares de nervios espinales
entran y salen de la médula a través de espacios entre las vértebras.
Los cuerpos celulares de las neuronas sensoriales están en los ganglios de la raíz dorsal por fuera de la médula espinal, y las fibras sensoriales llegan al lado dorsal de la médula espinal -en donde pueden establecer sinapsis con neuronas de proyección, interneuronas o neuronas motoras- o bien ascender hacia el cerebro. Las fibras de las neuronas motoras emergen de la zona ventral de la médula espinal. Los cuerpos celulares de las neuronas motoras localizados en la médula espinal pueden recibir señales de neuronas de proyección, de interneuronas y de neuronas sensoriales. Los cuatro tipos de neuronas frecuentemente están interconectadas en los arcos reflejos.
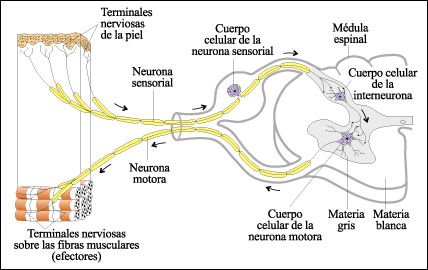
Un arco reflejo polisináptico.
Las terminales nerviosas libres
de la piel, cuando se estimulan de manera apropiada, transmiten señales
a lo largo de la neurona sensorial a una interneurona en la médula espinal.
La interneurona transmite la señal a una neurona motora. En consecuencia,
las fibras musculares se contraen.Las
neuronas de proyección, que no se muestran aquí, también
son estimuladas por la neurona sensorial y llevan la información sensorial
al cerebro.
El sistema nervioso somático
se divide en "voluntario" -controla los músculos esqueléticos
que pueden moverse a voluntad- e "involuntario" -incluye los nervios
motores que controlan al músculo cardíaco, las glándulas
y el músculo liso-.
Anatómicamente, las neuronas
motoras del sistema somático son distintas y están separadas de
las del sistema nervioso autónomo, aunque los axones de ambos tipos pueden
ser llevados dentro del mismo nervio.
Las divisiones del sistema nervioso
autónomo -simpática y parasimpática- son anatómica,
fisiológica y funcionalmente distintas. Funcionalmente, los sistemas
simpático y parasimpático son generalmente antagónicos.
La mayoría de los órganos internos están inervados por
axones de ambos sistemas y la regulación homeostática del cuerpo
depende de la cooperación de estas divisiones del sistema autónomo
y de la actividad de las glándulas endocrinas. El sistema parasimpático
está involucrado primariamente en las actividades restauradoras del cuerpo.
La estimulación parasimpática
hace más lenta la frecuencia cardíaca, incrementa los movimientos
del músculo liso de la pared intestinal, y estimula la secreción
de las glándulas salivales y de las glándulas digestivas del estómago.
El sistema simpático, por el contrario, prepara el cuerpo para la acción.
Los rasgos físicos del miedo, como el aumento de las frecuencias cardíaca
y respiratoria, entre otros, resultan de la descarga aumentada de neuronas del
sistema simpático.
El impulso nervioso
La conducción nerviosa
está asociada con fenómenos eléctricos. La diferencia en
la cantidad de carga eléctrica entre una región de carga positiva
y una región de carga negativa se llama potencial eléctrico. Casi
todas las membranas plasmáticas tienen una diferencia de potencial eléctrico
-el potencial de membrana- en el que el lado interno de la membrana es negativo
respecto al lado externo.
La transmisión del impulso nervioso es diferente de una corriente eléctrica: el impulso nervioso no experimenta disminución entre los extremos del axón; es mucho más lento que una corriente eléctrica y, a diferencia de ésta, la intensidad del impulso siempre es la misma: o bien no hay impulso nervioso en respuesta a un estímulo de una fibra nerviosa, o hay una respuesta máxima.
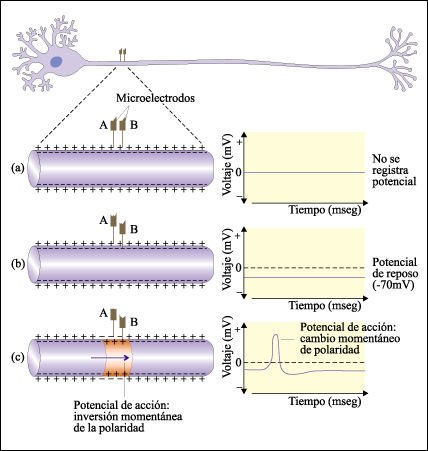
El potencial eléctrico a través de la membrana del axón se mide con microelectrodos conectados a un osciloscopio.
a) Cuando ambos electrodos están fuera de la membrana, no se registra
ninguna diferencia de potencial. b) Cuando un electrodo se coloca dentro de
la membrana, el interior de la neurona es negativo con respecto al exterior
y la diferencia entre los dos es de aproximadamente 70 milivoltios. Este es
el potencial de reposo. c) Al estimular un axón, el impulso nervioso
se propaga a lo largo de él; cuando alcanza la región en donde
se encuentran los microelectrodos, el osciloscopio muestra una breve inversión
de la polaridad: el interior se hace positivo en relación con el exterior.
Esta breve inversión en la polaridad es el potencial de acción.
El interior de la membrana está
cargado negativamente con respecto al exterior. Esta diferencia de voltaje -
la diferencia de potencial- constituye el llamado potencial de reposo de la
membrana. Cuando el axón es estimulado, el interior se carga positivamente
con relación al exterior. Esta inversión de la polaridad se denomina
potencial de acción. El potencial de acción que viaja a lo largo
de la membrana constituye el impulso nervioso.
Los potenciales de acción
registrados para una misma neurona casi siempre son iguales. La única
variación -aunque crítica- es la frecuencia, es decir, el número
de impulsos nerviosos que se producen en un tiempo determinado; la frecuencia
es directamente proporcional a la intensidad del estímulo.
Además de la medición
de la actividad de las neuronas, actualmente es posible registrar, en forma
macroscópica, la actividad cerebral por métodos no invasivos,
por medio de técnicas como la tomografía por emisión de
positrones o la resonancia magnética funcional que permiten determinar
qué zonas del cerebro están más o menos activas en base
a cambios en distintos parámetros fisiológicos cuando se realizan
tareas específicas (sensoriales, motoras o cognitivas). Esta actividad
general se correlaciona con la de las neuronas individuales, la cual es determinada
con microelectrodos colocados a ambos lados de la membrana neuronal.
El potencial de acción
depende del potencial eléctrico neuronal, que, a su vez, es posible por
las diferencias en la concentración iónica a cada lado de la membrana.
En los axones, las diferencias críticas de concentración involucran
iones potasio (K+) e iones sodio (Na+).
La distribución de los
iones a ambos lados de la membrana es característica y es gobernada por
tres factores: 1) la difusión de partículas a favor de un gradiente
de concentración, 2) la atracción de partículas con cargas
opuestas y la repulsión de partículas con cargas iguales y 3)
las propiedades de la propia membrana.
La bicapa lipídica de la membrana del axón es impermeable a los iones y a la mayoría de las moléculas polares, por lo que el movimiento de partículas a través de la membrana depende de proteínas que proporcionan canales que las partículas pueden atravesar por difusión facilitada o por transporte activo. Los iones son específicos, particularmente Na+ y K+. Otro rasgo significativo de la membrana del axón es la presencia de una proteína integral de membrana -la bomba de sodio-potasio- que bombea iones Na+ hacia afuera del axón e iones K+ hacia adentro.
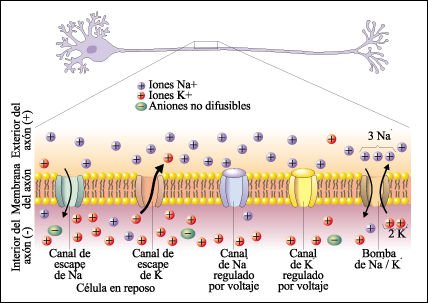
Axón en estado de reposo.
En la bicapa lipídica del axón hay proteínas integrales
de membrana que actúan como canales, por los que los iones K+ y Na+ se
pueden desplazar entre el citosol de los axones y el líquido intersticial
externo. Los distintos tipos de canales son el canal de escape de Na+ y el canal
de escape de K+ permanecen siempre abiertos, y durante el estado de reposo permiten
la difusión de los iones hacia adentro y hacia fuera del axón
siguiendo su gradiente de concentración. Los canales de Na+ y los canales
de K+ regulados por voltaje permanecen cerrados durante el estado de reposo.
La bomba Na+/ K+ bombea 3 iones Na+ hacia fuera del axón por cada 2 iones
K+ bombeados hacia adentro. La concentración de iones K+ es mucho mayor
en el citosol que en el líquido intersticial. Por lo tanto, los iones
K+ difunden hacia fuera del axón a través de los canales de escape
de K+, a favor de su gradiente de concentración. Los iones más
grandes, cargados negativamente, no pueden acompañar a los iones K+ en
su camino hacia fuera del axón. En consecuencia, el interior del axón
se carga negativamente en relación al exterior. La bomba Na+/ K+ extrae
rápidamente iones Na+ del axón, a la vez que aumenta la concentración
de iones K+ por el bombeo de esos iones hacia el interior. Con ello se mantienen
las diferencias de concentración de las que depende el potencial de la
célula en reposo.
La membrana axónica está polarizada, el interior es más negativo que el exterior, lo que determina el potencial de reposo. Esto es lo que hace posible la generación de un potencial de acción. La carga negativa en el interior del axón atrae un cierto número de iones K+ y Na+ que se dirigen hacia el interior del axón por sus respectivos canales de escape. Los iones Na+ se extraen rápidamente del axón gracias a la bomba Na+/ K+, a la vez que aumenta la concentración de iones K+ por el bombeo de esos iones. Con ello se mantienen las diferencias de concentración de las que depende el potencial de la célula en reposo.
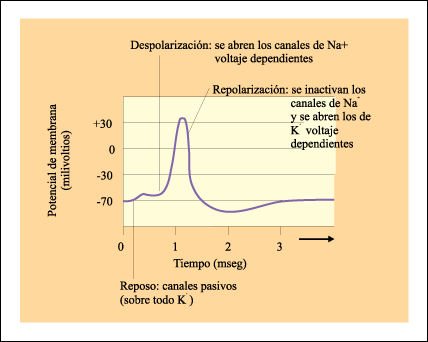
El potencial de acción.
Una porción de la membrana se vuelve momentáneamente muy permeable
a los iones Na+ mediante la apertura de canales dependientes de voltaje. Cuando
se abren los canales de Na+, los iones pasan precipitadamente al interior y
la polaridad de la membrana se invierte. A continuación, aunque no se
observa en esta figura, se abren los canales de potasio regulados por voltaje
y se inactivan los de sodio regulados por voltaje, lo que lleva a una repolarización
de la membrana y al eventual reestablecimiento del estado de reposo. b) Gráfico
de un potencial de acción y los movimientos iónicos asociados.
Un aspecto importante del impulso
nervioso es que, una vez iniciado, la inversión transitoria de la polaridad
continúa moviéndose a lo largo del axón, renovándose
continuamente.
El potencial de acción
se autopropaga porque en su pico, cuando el interior de la membrana en la región
activa es comparativamente positivo, los iones cargados positivamente se mueven
desde esta región al área adyacente dentro del axón, que
todavía es comparativamente negativa. Como resultado, el área
adyacente se despolariza o, sea, se hace menos negativa. Esta despolarización
abre los canales de Na+ activos y regulados por voltaje, que permiten que los
iones Na+ entren precipitadamente. El incremento resultante en la concentración
interna de iones Na+ despolariza la siguiente área contigua de la membrana,
haciendo que sus canales iónicos de Na+ se abran y permitiendo que el
proceso se repita. Como consecuencia de este proceso de renovación, que
se repite a lo largo de toda la membrana, el axón -un conductor muy pobre
de la corriente eléctrica- es capaz de conducir un impulso nervioso a
una distancia considerable sin que cambie en absoluto la intensidad. El impulso
nervioso se mueve en una sola dirección porque el segmento del axón
situado "detrás" del sitio donde se produjo el potencial de
acción tiene un período refractario breve durante el cual sus
canales iónicos de Na+ no se abrirán; así, el potencial
de acción no puede retroceder.
Los axones largos de los vertebrados generalmente están envueltos en vainas de mielina, formadas par células de la glia especializadas. La vaina de mielina hace que la propagación del impulso nervioso sea mucho más rápida en los vertebrados que en los invertebrados.
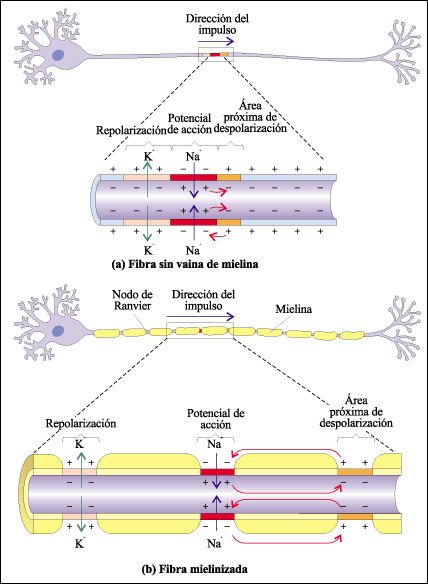
Fibras con y sin vaina de mielina.
a) En una fibra sin vaina de
mielina, toda la membrana del axón está en contacto con el líquido
intersticial. Todas las partes de la membrana contienen canales y bombas de
sodio-potasio. b) En una fibra mielinizada, en cambio, solo están en
contacto con el líquido intersticial las zonas de la membrana axónica
correspondientes a los nodos de Ranvier. Prácticamente todos los canales
iónicos y bombas de sodio-potasio se concentran en estas zonas. Así,
los potenciales de acción se pueden generar solo en los nodos y el impulso
nervioso salta de nodo en nodo, acelerándose la conducción.
Las sinapsis
Las señales viajan de
una neurona a otra a lo largo de la unión especializada -la sinapsis-
que puede ser de naturaleza química o eléctrica.
La llegada de un potencial de
acción a la terminal axónica de la célula presináptica
está acompañada por cambios en la concentración iónica.
Estos cambios son transmitidos a través de las uniones nexus a la célula
postsináptica, donde despolarizan la membrana celular e inician un nuevo
potencial de acción.
Una sinapsis química.
La llegada de un potencial de acción en la terminal axónica inicia
la fusión de vesículas sinápticas con la membrana del axón,
liberando neurotransmisores en el espacio sináptico. Éstos difunden
a la célula postsináptica, donde se combinan con receptores específicos
de la membrana celular. Una red proteica en el espacio sináptico ancla
a las membranas presinápticas y postsinápticas y, en ocasiones,
contiene enzimas que degradan las moléculas de neurotransmisor.
Algunos neurotransmisores son
sintetizados en el cuerpo celular de la neurona y transportados a los terminales
axónicos, donde son "empaquetados" y almacenados en vesículas
sinápticas. Otros son sintetizados y se empaquetan dentro de las terminales
axónicas. La liberación de las moléculas neurotransmisoras
es disparada por la llegada de un potencial de acción al terminal axónico.
Después de su liberación, los neurotransmisores son removidos
o destruidos rápidamente, interrumpiéndose su efecto; ésta
es una característica esencial del control de las actividades del sistema
nervioso.
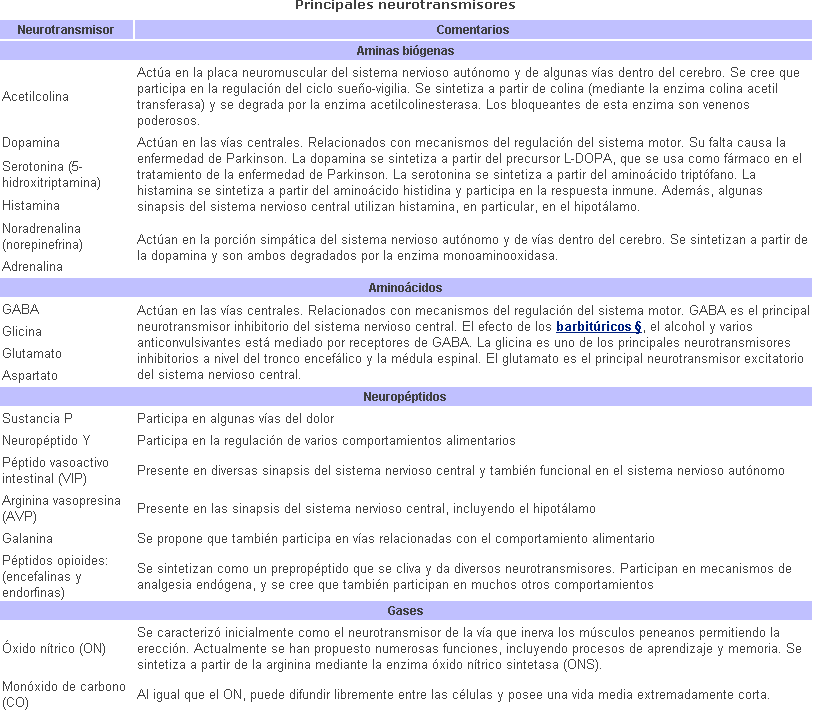
Una variedad de sustancias químicas
funcionan como neurotransmisores. En el sistema nervioso periférico,
los principales son la acetilcolina y la noradrenalina.
En el sistema nervioso central
se han encontrado muchos otros neurotransmisores, incluyendo a las llamadas
aminas biógenas (como la noradrenalina) entre ellas la dopamina y la
serotonina, ambas derivadas de aminoácidos.
Casi todas las drogas que actúan
en el cerebro alterando el humor o el comportamiento, lo hacen intensificando
o inhibiendo la actividad de los sistemas neurotransmisores. La cafeína,
la nicotina y las anfetaminas, estimulan la actividad cerebral en forma análoga
a los neurotransmisores excitatorios en las sinapsis. La cloropromazina y los
tranquilizantes relacionados bloquean los receptores de dopamina en muchos sitios,
mientras que el ácido lisérgico -LSD- (un alucinógeno)
inhibe la acción de la serotonina cerebral.
Debe mencionarse que muchos
neuropéptidos, junto con otras sustancias neuroactivas, pueden desempeñar
otro papel en la transmisión sináptica; no generar la señal
transmisora sino regularla. Estas moléculas, que pueden ser liberadas
de las mismas terminales axónicas que los neurotransmisores principales
o de otras células, se conocen como neuromoduladores.
Aunque éstos pueden moverse
directamente a través de la hendidura sináptica, también
pueden difundir a una distancia mayor, afectando a numerosas células
dentro de una región local del sistema nervioso central. Al igual que
los neurotransmisores, se unen a receptores específicos de membrana y
alteran los canales iónicos o ponen en movimiento segundos mensajeros;
sus efectos frecuentemente consisten en modular la respuesta de la célula
a un neurotransmisor principal. Se han identificado hasta el momento más
de 200 sustancias diferentes que funcionan como neuromoduladores. Estas incluyen
las endorfinas, los interferones y las interleucinas, las hormonas liberadoras
hipotalámicas, las hormonas hipofisarias, las hormonas de páncreas
como la insulina, y hasta las hormonas digestivas gastrina y colecistocinina.
Las dendritas y el cuerpo celular
de una sola neurona pueden recibir señales -en forma de moléculas
de neurotransmisor o neuromodulador- enviadas por centenares o hasta por miles
de sinapsis. La unión de cada molécula a su receptor tiene cierto
efecto en el grado de polarización de la célula postsináptica.
Si el efecto es que el interior de la célula se vuelve menos negativo
(despolarización) se dice que es excitatorio. Por el contrario, si el
efecto es que se mantiene al potencial de membrana en valores cercanos al potencial
de reposo, o aun, el interior se hace más negativo (hiperpolarización),
se dice que es inhibitorio.
Los cambios en la polaridad
inducidos por los neurotransmisores y los neuromoduladores se extienden desde
las sinapsis a través de la célula postsináptica al cono
axónico, que es la región del axón en la cual puede originarse
un impulso nervios. Si el efecto colectivo es una despolarización suficiente
como para permitir un flujo de iones Na+ tal que constituya el inicio de un
potencial de acción, entonces comienza un impulso nervioso en el axón
de la célula postsináptica y un nuevo mensaje es enviado velozmente
a una multitud de otras neuronas con las cuales hace sinapsis el axón.
El procesamiento de la información
que ocurre dentro del cuerpo celular de cada neurona individual desempeña
un papel central en la integración y en el control ejercidos de manera
conjunta por los sistemas nervioso y endocrino. Es afectado no sólo por
los neurotransmisores y neuromoduladores específícos recibidos
por la célula, sino también por su cantidad, el tiempo preciso
de su llegada y las localizaciones en la neurona de las varias sinapsis y receptores.


